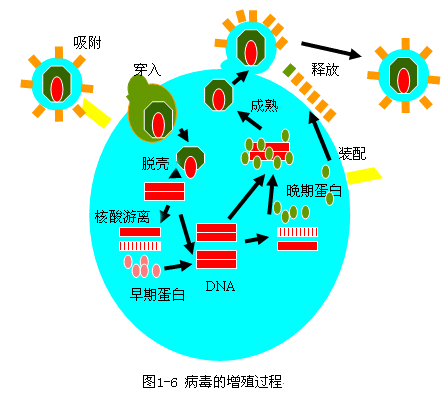
|
二、病毒的生物学特性
1、大部分病毒没有酶或酶系统极不完全,不含催化能量代谢的酶,不能进行独立的代谢功能。
2、严格的活细胞内寄生,没有自身的核糖体,不能生长也不进行二均分裂,必须依赖宿主细胞进行自身的核酸复制,形成子代。
3、在寄主细胞内的病毒对各种化学药剂和抗菌素不敏感。
三、病毒的理化特性
病毒受理化因素作用后失去传染性,称为灭活。
1.物理因素
 温度 病毒大多数耐冷不耐热,除肝炎病毒外,多数病毒加热56℃30min或100℃几秒钟即可灭活。室温下短时间死亡。在0℃以下温度生存良好。 温度 病毒大多数耐冷不耐热,除肝炎病毒外,多数病毒加热56℃30min或100℃几秒钟即可灭活。室温下短时间死亡。在0℃以下温度生存良好。
辐射 病毒对射线敏感。电离辐射中的χ线和γ射线使核苷酸链发生断裂,而紫外线照射可使核苷酸链形成胸腺嘧啶二聚体,抑制病毒核酸的复制。
2.化学因素
脂溶剂 包膜病毒对脂溶剂敏感,因其包膜富含脂类,因而易被乙醚、丙酮、氯仿、阴离子去垢剂等脂溶剂所溶解,使病毒失去感染的能力,无包膜病毒对脂溶剂有抗性。
酚类 可使病毒蛋白质变性,去除病毒的衣壳蛋白或破坏包膜病毒的脂蛋白膜,故可作为病毒的消毒剂。常与去垢剂合用(如1%~5%苯酚)可使许多病毒灭活。
醛类 甲醛对病毒蛋白质和核酸都有破坏作用,使病毒失去感染性,是常用的灭活剂。甲醛也可与蛋白质氨基酸发生反应,但对蛋白质的构型作用不强,因此对免疫原性影响不大,故常用于病毒灭活疫苗的制备。
氧化剂、卤素及其化合物 病毒对过氧化氢、漂白粉、高锰酸钾、碘和碘化物及其它卤素类化学物质都很敏感,为有效的病毒灭活剂。70%甲醇及乙醇能使大多数病毒灭活。次氯酸盐、过氧乙酸等对肝炎病毒等有较好的消毒作用。
四、病毒的传播途径
1、水平传播
水平传播是病毒在人群个体之间的传播,也包括由媒介动物参与的传播,此传播途径主要是经皮肤和粘膜表面感染。
经皮肤传播 有些病毒通过注射、输血或机械性损伤从皮肤侵入机体而致病,如乙型肝炎病毒、人类免疫缺陷病毒等;有些通过昆虫叮咬、动物咬伤等方式进入宿主体内而致病,如流行性乙型脑炎病毒、狂犬病病毒。
经粘膜表面传播 粘膜表面的上皮细胞是许多病毒最先侵入并增殖的部位。多数病毒可经呼吸道、消化道粘膜侵入机体,如流感病毒、脊髓灰质炎病毒等;少数病毒可通过接触传播,如直接接触眼结膜引起角膜结膜炎,通过性接触,病毒经泌尿生殖道粘膜感染引起性病的传播。有些病毒感染仅局限于呼吸道或消化道粘膜,如流感病毒、轮状病毒等;有些病毒经粘膜表面感染后,通过淋巴或血流扩散至其它器官,引起病毒血症或相应器官的典型表现,如麻疹病毒、脊髓灰质炎病毒等。
2、垂直传播
通过胎盘、产道及哺乳,病毒直接由亲代传给子代的感染方式称为垂直传播。病毒可通过胎盘传给胎儿或经产道、哺乳传给新生儿,是病毒感染的特点之一,其它微生物是极少见的。孕妇感染某些病毒后,尤其在妊娠三个月以内时,易经胎盘传给胎儿。现在已知十余种可经垂直传播的病毒,其中以风疹病毒、乙型肝炎病毒、巨细胞病毒及人类免疫缺陷病毒为多见,可引起早产、死胎或先天畸形等。
|

 返回
返回